これまで「中国の世界観」「西アジアの世界観」と、自然外圧や同類圧力が全く異なる集団群の世界観を追求してきました。
そして、今回は「日本の世界観」に迫っていきます。日本の世界観の出発点とも言える「縄文時代」に形成された世界観とは、どのようなものであったのでしょうか。
■自然を対象に、追求して、一体感を高め、世界観として共有してきた日本人

こちらから引用させていただきました
日本は豊かな森林や海・河川があり、また、温暖湿潤な気候で、これまでに追求してきた中国や西アジアのように生きていくための場所が限られてはいません。
なので、世界観の全く異なる集団同士が出会い、再統合に迫られるということは多くなかったと思われ、基本的には、単一の集団内で世界観が塗り重ねられ、連綿と引き継がれていったものと考えられます。
再統合しなくてはならないという同類圧力は中国や西アジアに比べると低いものの、常に山、川、海、草木、動物に囲まれて生きており、また、地震・火山噴火・津波・河川氾濫などの突発的で圧倒的なエネルギーに飲み込まれることもあり、もっぱら自然を対象に、みんなでとことん追求して、一体感を高め、世界観として共有していたのでしょう。
その追求は、現代にまで、残され、引き継がれてきたものから窺い知ることができます。
例えば、、、
・土器・土偶 ・環状列石 ・縄(精霊信仰)
・発酵・保存技術 ・大和言葉
・海洋技術 ・建築技術
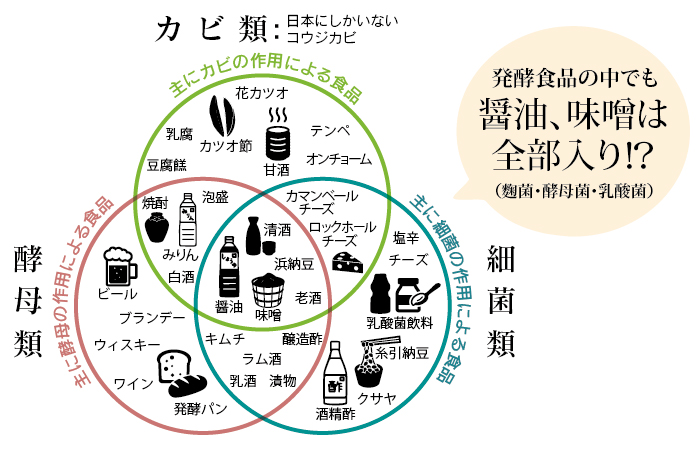
などは、縄文時代を表す特徴的な追求です。早速、日本が世界に誇る技術・文化の一つである「発酵」から日本の世界観を捉えてみたいと思います。